

In an ideal situation the COVID-19 pandemic would be brought under control by vaccine-induced immune responses in most individuals. The reality that might be achieved is a combination of vaccine-induced and some natural convalescent immunity. Studies to date indicate that the two are compatible and relatively comparable (Carvalho et al., 2021; Stamatatos et al., 2021). Since the publication of our mRNA vaccines blog this past January and the viral vector vaccines blog in April, concerns related to changes to the SARS-CoV-2 genome have become increasingly urgent. Due to the large number of people who have become infected worldwide and the measurable rate of replication errors by viral RNA-dependent RNA polymerase (RdRp) there are now many occurrences of mutations in the virus at large. Indeed, a recent review quoted that with the known rates of evolution in the spike protein gene, the mutation rate (~10-6 mutations/site/replication cycle) is high enough to mutate on average every amino acid codon in the spike encoding gene at least once per infected person (Winger and Caspari, 2021)!
There is intense activity and attention now focused on whether and how mutations in the SARS-CoV-2 genome will affect both natural and vaccine-induced immunity (Plante et al., 2021). The immune system correlates of protection are still not fully defined (Dan et al., 2021, Stamatatos et al., 2021) but clearly, there is some correlation between neutralizing antibody (nAb) response and protection (Carvalho et al., 2021). It is also clear that CD4+ an CD8+ T cell responses are strong amongst many infected individuals, and this component of the immune response follows its own kinetics (Dan et al., 2021). There is also evidence that contributions by CD4 and CD8 are important for maintenance of immunity, but the science is still unclear how these mesh with humoral immunity to create full correlates of protection.
After infection, the majority of nAbs responses in convalescent serum is observed against the spike protein, the main reason why most vaccine preparations target the spike component of the virus. Among the nAbs to spike protein in sera most react to sites in the receptor-binding domain (RBD) (Cele et al., 2021, McCallum et al., 2021). Almost all of the remaining nAb responses are to the N-terminal domain (NTD). Observed mutations in the spike protein are generally driven either by increased viral fitness, host adaptation, or immune escape pressure (Plante et al., 2021). In this article, we will discuss some of the mutations that occur in the spike protein and are of highest concern, and how these mutations are occurring together in viral lineages/variants. Lastly, we will review our emerging understanding of how well existing vaccines perform against these variants.
Critical SARS-CoV-2 Surface Components
Briefly, the SARS-CoV-2 S protein is comprised of an N-terminal S1 subunit that is responsible for receptor binding and a C-terminal S2 subunit that mediates virus fusion to the cell membrane (McCallum et al., 2021). The S1 subunit is further described as containing an N-terminal domain (NTD) and the receptor-binding domain (RBD). The majority of nAb responses in convalescent sera are directed against the RBD and in some individuals constitute more than 90% of total neutralizing activity (Greaney et al., 2021). Within the RBD exists the receptor-binding motif (RBM) which is the part of the receptor that directly contacts the host angiotensin-converting enzyme 2 (ACE2) embedded in the host cell membrane. It is important to point out that not all antibodies that bind to the RBD bind to sites within the RBM but the RBM-binding nAbs tend to be most potently neutralizing (Greaney et al., 2021) and often compete for binding with ACE2. Nevertheless, there are also significant contributions to neutralizing activity mediated by nAbs that recognize sites in the NTD. Within the NTD is an antigenic super site that is recognized by some very potent nAbs isolated from sera (McCallum et al., 2021).
Within this combined landscape of known sites of high antigenicity there have been numerous descriptions of mutations occurring in the population at large that have been reported by multiple labs and across many countries.
Mutations in the SARS-CoV-2 Genome
Interestingly, the numerous mutations that have been observed so far affect both the viral activity and the response by the immune system. Here, we will detail a few of the most critical mutations.
- D614G: A mutation that was discovered early in the pandemic. D614G has since become predominant and appears in sequences from infected individuals across the world. It appears that D614G enhances virus biding to ACE2, as well as increasing transmissibility and viral replication (McCallum et al., 2021). Despite the fact that D614 is outside the RBD its occurrence promotes opening (or up configuration) of more than one RBD per spike trimer (Winger and Caspari, 2021). However, it appears D614G alone is not enough to cause immune evasion from any of the vaccines in current use (Thompson et al., 2021).
- N439K: A more recently emerging mutation is the N439K mutation (Thompson et al., 2021). As of January 6, 2021, the N439K mutation was the second most commonly observed receptor binding domain (RBD) mutation worldwide. N439K results in enhanced RBD affinity to the ACE2 receptor but does not result in much change in the clinical spectrum. However, the mutation does result in immune escape from some polyclonal sera and monoclonal antibody (mAbs).
- N501Y: A very interesting mutation found to be occurring across the globe in multiple viral lineages. The N501Y mutation has arisen independently multiple times and is shared by three significant lineages B1.351, B.1.1.7, and P.1, all of which we will discuss in depth later in this report. N501 is near the C-terminal boundary of the RBD and part of the ACE2 interaction surface (Zhou et al., 2021). N501Y apparently contributes both to increased affinity of the RBD to ACE2 and likely increased transmissibility for viruses that contain it.
Multiple groups are working to understand the broader landscape of mutations in the spike glycoprotein that may be immune escaping due to selection in the population. One group using a chimeric virus model system has uncovered upwards of 50 escape mutants in the RBD (Liu, Z., et al., 2021).
- Two significant mutants S477N and E484K that were described by Liu, Z., et al. (2021), that allow immune escape in cell culture, have been seen in viral sequences worldwide. Another group working with a yeast-display model system also confirmed E484 as an important mutation target site (Greaney et al., 2021). This group found that E484 mutations can reduce neutralization by plasma from recovered COVID-19 patients more than 10-fold.
- K417 is another site in the RBD that makes contact with ACE2 and is prone to mutations with effects on Ab neutralization (Greaney et al., 2021). K417 mutations to N or T residues have shown up in viral sequences obtained from clinical isolates. Evidence suggests that mutations like K417N/T help lock the spike protein structure into the ‘one-RBD-up’ conformation potentially contributing to both immune escape (Zhou et al., 2021, Wang et al., 2021) and better ACE2 binding (Wang et al., 2021b). However there is also data indicating that K417N mutations may on their own, by increasing the open ACE2-binding confirmation, expose other epitopes for nAb activity (Li et al., 2021)
- NTD mutations: There are a number of mutations in the NTD that have been noted in genomic sequences of SARS-CoV-2 worldwide. Some of the most prevalent that are associated with changes in Ab neutralization are L18F, T19A, ∆69-70, D80A, Y144 deletion, D215G, ∆242-244, R426A (McCallum et al., 2021). In particular the NTD region is the focus of deletion mutations that drive antibody escape (McCarthy et al., 2021). Some of mutations in the NTD occur in combination with other mutations in variants or lineages (see later in this report).

Figure 1: The receptor binding motif of spike (RBD) with a view into the binding site in (a), and the backside of the binding site in (b). Some of the key amino acids mutated in the variants are highlighted (image source Winger and Caspari (2021).
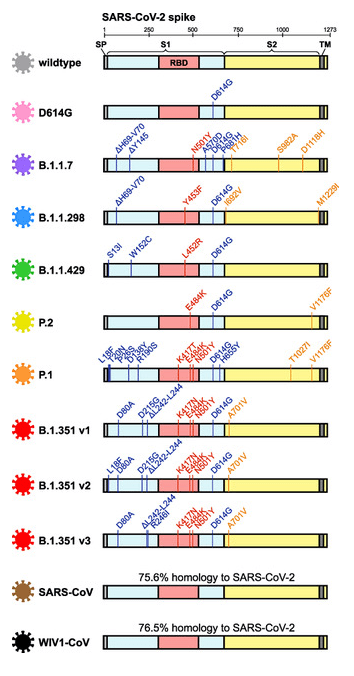
Figure 2:S Schematic summarization of SARS-CoV-2 mutations in the spike protein sequence of SARS-CoV-2 variants (image source Garcia-Beltran et al., 2021).
Of course, the examples provided only scratch the surface of current work on individual spike protein mutations and their effects. Table 1 summarizes spike protein mutations visualized in Figure 1 and 2.
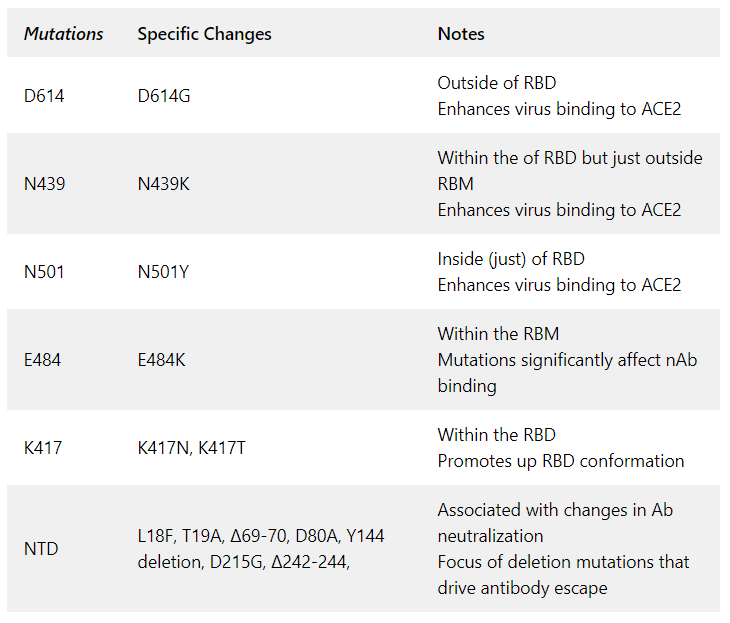
Table 1: Summary of spike protein mutations.
Discovery and description of variants and lineages
Single point mutations in spike glycoprotein or other SARS-CoV-2 proteins are concerning enough, but serious attention is now focused on virus variants or lineages (see weekly epidemiology summaries from WHO). These are cases where multiple mutations have accumulated in evolving viral strains. The WHO labels the most important and potentially dangerous as Variants of Concern (VOC) as compared to Variants of Interest (VOI) that are currently just monitored. It should be noted that SARS-CoV-2 variants and lineages are defined by mutations carried in the entire viral genome but we will confine our discussions to the defining mutations in spike.
The efforts to identify new mutations and variants and monitor the rates of change in existing ones is hampered by the uneven SARS-CoV-2 genomic surveillance performed worldwide via viral DNA sequencing. Another complication delineated just at the beginning of June 2021, is that two commonly used protocols for SARS-CoV-2 genomic sequencing can be compromised by the mutations which disrupt the function of sequencing primers (Kuchinski et al., 2021). To simplify the naming of the many variants evolving across the globe and partially driven by the desire to have a set of less politically-sensitive names, the WHO announced on May 31, 2021 simple, easy-to-say labels for SARS-CoV-2 VOCs and VOIs. Table 2 summarizes the various variants and their region/country of origin.
| WHO Name | Variant | Scientific Name |
| Alpha | Kent, U.K. | B.1.1.7 (aka 501Y.V1 or 20I) |
| Beta | South Africa | B.1.351 (aka 501Y.V2 or 20H) |
| Gamma | Brazil | P.1 (aka 501Y.V3 or 20J) |
| Delta | India | B.1.617.2 |
- B.1.351 (aka 501Y.V2 or 20H or WHO Beta) – South African variant: Not the first to be recognized by the scientific community, but one of the most concerning of all variants defined so far is the B.1.351 variant (Tegally et al., 2021). B.1.351 was first recognized in late 2020 and modeled to have emerged in summer 2020 in South Africa, hence it being commonly referred to as the “South Africa variant.” This lineage is defined in part by spike point mutations L18F, D80A, D215G, R446I, A701V, one deletion ∆242-244 and a triplet of mutations in the RBD; K417N, E484K and N501Y (Garcia-Beltran et al., 2021). The triple occurrence of mutations in the RBD is particularly concerning as these mutations combine to both increase affinity to ACE2 and avoid neutralization by antibodies. Once B.1.351 emerged, it spread rapidly and became the dominant variant in South Africa by displacing other lineages, presumably due to selection advantage in transmissibility, immune evasion, or both (Tegally et al., 2021). Since then, B.1.351 has gone on to be detected in many countries all over the world.
- B.1.1.7 (aka 501Y.V1 or 20I or WHO Alpha) – Kent, U.K. variant: Another lineage that has garnered much attention is B.1.1.7, colloquially known as the “UK variant”. This variant shares the N501Y and D614G mutations with B.1.351. There is considerable data about B.1.1.7, mostly because the UK viral sequence surveillance campaign is quite robust (COVID-19 Genomics UK Consortium) compared to much of the rest of the developed world. The B.1.1.7 lineage was spotted early in September 2020 in England and spread rapidly across the country to become the dominant lineage (Washington et al., 2021). B.1.1.7 contains the signature spike protein mutations ∆69-70, Y144 deletion, A570D, P681H, T716I, S982A, D1118H as well as the aforementioned critical RBD mutations N501Y. To-date, B.1.1.7 has been detected in over 75 countries around the world and has been rapidly expanding in the U.S. since November 2020 (Washington et al., 2021). There is evidence that B.1.1.7 is both more transmissible and may lead to higher mortality than ancestral strains.
- P.1 (aka 501Y.V3 or 20J or WHO Gamma) – Brazil variant: A third widely discussed and concerning lineage is the P.1 variant, often referred to as the “Brazil variant”. Viral genome sequencing and modeling indicates P.1 arose in Manaus around November 2020 (Faria et al., 2021). P.1 contains the ten signature spike protein mutations L18F, T20N, P26S, D138Y, R190S, K417T, E484K, N501Y, H655Y and T1027I compared with its immediate ancestor lineage B.1.1.28. P.1 has the same three core residues mutated as B.1.351, but interestingly, since having arisen independently indicates the convergent nature of adaptation in the RBD. Cryo-EM structural modeling shows the P.1 spike trimer is exclusively in the one-RBD-up confirmation (Wang et al., 2021b). Early data indicates that P.1 is more transmissible by a median of 2 times and possibly more lethal as well though that has not been confirmed.
- B.1.617.2 (aka WHO Delta) – Indian variant: The lineage that has been contributing to the very concerning situation in India is B.1.617.2. Initial descriptions of B.1.617 as a VOC were later followed by recognition of three subtypes B.1.617.1, B.1.617.2 and B.1.617.3, with B.1.617.2 (Delta) variant being deemed the most concerning (WHO update 42). B.1.617.2 contains 12 mutations in the spike protein compared to the original Wuhan isolate including the now ubiquitous D614G mutation. The other defining spike protein mutations are T19R, K77R, G142D, ∆156-157, R158G, A222V, L452R, T478K, P681R and D950N (Wall et al., 2021). B.1.617.2 does contain two mutations within the RBD, L452R, T478K, but none that overlap with the other major VOCs described above. The T478K mutation lies adjacent to the S477N mutation identified in mutational studies and is close to E484 in 3-D space. It is therefore suggested to play a role similar to E484K (Planas et al., 2021b). It also has several mutations and deletions in the NTD which might disrupt antibody binding to the critical supersite. B.1.617.2 is believed to have increased transmissibility as delineated in a study from authors at Public Health England (Allen et al., 2021). A measure of concern is the rapidity with which the information about this variant is evolving. A newer sub-lineage of the Delta variant was first reported on June 11, 2021 by Public Health England (now called B.1.617.2.1 or Delta plus or AY.1) carrying the K417N mutation has by now been sequenced in samples from at least 11 countries.
- B.1.427 and B.1.429 (aka WHO Epsilon) – Cal variant: Another lineage, found to be originating in the continental U.S., includes the two closely related B.1.427 and B.1.429 variants. Both variants were seen the first time in California in May of 2020. The B.1.427 and B1.429 variants share a trio of characteristic mutations, S13I, W152C, and L452R in the spike protein (Deng et al., 2021). Note some sources list B.1.427 as only containing L452R. By early 2021, this variant has been accounted for over 50% of all observed cases in California. The two variants, B.1.427 and B1.429, together have been associated with somewhat increased transmission risk, but not a definite worse prognosis (Deng et al., 2021),
- Other VOI: There are currently four additional VOI, designated as Eta through Lambda (Zeta having been downgraded) by the WHO, found in many different regions. Data on these variants is mixed due to uneven depth of sequence surveillance coverage which is unfortunate and potentially hazardous to the goal of defeating the pandemic.
Concerns about Vaccine Performance, Monoclonal Antibody Activity, and Natural Immunity Against SARS-CoV-2 Variants
Mutations in the spike protein are present at large in very significant numbers and with significant diversity. The question naturally arises: what does this mean for the pandemic, especially with regards to disease treatment, vaccine performance, and convalescence of individuals? There is already significant and rapidly accumulating data, for a number of variants, concerning how effectively previously infected patient sera collected after convalescence maintain the ability to neutralize various SARS-CoV-2 variants in culture. Likewise, there is considerable information on how the sera of vaccinated individuals treated with one of the mRNA vector or viral vector vaccines retain its neutralization potential against various variants. In addition, there is considerable information about how various monoclonal antibody drugs retain activity against or fail to neutralize variants in cell culture models. There is some, but still far more limited information about performance of the vaccines against various lineages in the population at large. We will do our best to present some key findings from this rapidly changing bolus of information, and this specifically around four key variants.
Vaccine Performance Against Specific SARS-CoV-2 Variants
- B.1.351 (Beta): Significant research focused on the B.1.351 variant after a relatively early report indicated near total failure of the AZD vaccine (AstraZeneca-University of Oxford) to protect people from this variant in a South Africa study (Madhi et al., 2021). Three key groups found that convalescent serum had reduced neutralization potential with some samples lacking any activity versus B.1.351 (Planas et al. 2021, (Liu, Y., et al., 2021, Wang et al., 2021). The same studies also found that both Moderna and Pfizer vaccinated individuals had reduced serum neutralizing activity by varying amounts for theB.1.351 (Beta) lineage. They also found dramatically reduced binding activity of some mAbs when used in treatment of COVID-19, mostly the ones that have biding sites on or near K417N or E484K mutations. Multiple other groups have found general agreement with these trends though the absolute numbers differ (Cele et al., 2021, Garcia-Beltran et al., 2021, Hoffmann et al., 2021, Li et al., 2021, Stamatatos et al., 2021, Wall et al., 2021, Zhou et al.,2021 and many others). When vaccine efficacy is studied in the context of the variants there is some indication that despite lower serum neutralization titer BNT162b2 (Pfizer-BioNTech) is still up to 75% effective against B.1.351 in Qatar (Abu-Rabbad et al. 2021). Current data supports the hypothesis that though B.1.351 is more dangerous in multiple ways the current vaccination regimens with RNA-based vaccines should still be effective in its presence but more studies are warranted for confirmation.
- B.1.1.7 (Alpha) : In part due to early acquisition of sequence data and also concerns over later waves of COVID-19 in the UK, there have been a considerable number of studies conducted that include the B.1.1.7variant. Multiple groups have found that B.1.1.7 has either modestly reduced or essentially unchanged neutralization when tested with either convalescent serum or vaccine serum (Planas et al. 2021, Wang et al. 2021, Garcia-Beltran et al., 2021, Hoffmann et al., 2021, Li t al., 2021, Wall et al., 2021). Wang et al. (2021) found B.1.1.7 led to substantial reduction in neutralizing activity of mAbs directed against the NTD but not generally to those directed at the RBD. Abu-Rabbad et al. (2021) found BNT162b2 (Pfizer-BioNTech) to be 87% effective versus B.1.1.7 at a time when B.1.1.7 and B.1.351 were together responsible for almost all community COVID-19 cases. Taken in total, this data suggests that though B.1.1.7 may pose a higher individual risk (via potentially worse clinical outcomes), it alone will likely not threaten the efforts to combat the pandemic through vaccination.
- P.1 (Gamma): The devastating COVID outbreak in Brazil has led to intense scrutiny of the P.1 lineage though the data are not yet as deep as for some other variants. Three major studies (Garcia-Beltran et al., 2021, Hoffmann et al., 2021, Wang et al., 2021b) agree that neutralization of P.1 was reduced (2.2-6.7 fold) for BNT162b2 vaccine serum, (2.8-4.8 fold) for mRNA-1273 vaccine serum and (3.4-6.4 fold) for convalescent serum. Generally the reductions seen with P.1 are intermediate between B.1.351 (Beta) and B.1.1.7 (Alpha). Hoffmann et al. (2021) found the P.1 variant to be completely resistant to some single therapeutic monoclonal antibodies while Wang et al. (2021b) found P.1 to be refractory to neutralization by 3 mAbs that already have EUA. There is currently no published evidence of the effectiveness of the major vaccines in use in populations where P.1 is present at higher level. However the fact that P.1 emerged in an area despite very high previous levels of infection with ancestor strains is concerning in itself (Faria et al., 2021).
- B.1.617.2: B.1.617 (Delta) is a more recently identified variant and as such much of the information currently published is based on non-peer reviewed data. Hoffman et al. (2021b) have reported that B.1.617 is resistant to neutralization by convalescent serum (2.0 fold) and BNT162b2 serum (3 fold). These reductions in neutralization are both significantly less compared to the reduction of B.1.1351 (Beta) when tested in parallel with the same assays. However this virus version had R21T, E154K, Q218H, L452R, E484Q, D614G and P681R mutations in spike. Planas et al. (2021b) later showed that B.1.617.2 was resistant to neutralization by convalescent serum (4-6 fold) similar to B.1.1351 when assayed in parallel. In the same study BNT162b2 serum neutralization was reduced (3-6 fold) but significantly less than B.1.1351 while recipients of a single dose of AZD1222 had barely any ability to neutralize the variant. Just at the beginning of June another group published that spike variants were neutralized by BNT162b2 serum with decreasing titer: wt > B.1.617.2 > B.1.618 > B.1.525 > B.1.617.1 (Liu, J., et al., 2021). One non-peer reviewed observational study based on public health data from the UK showed that the BNT162b2 vaccine efficacy was 88% with B.1.617.2 versus 93% with B.1.1.7 (Bernal et al., 2021). With AZD1222 effectiveness dropped from 66% to 60%. This study was based on multiple weeks of data wherein 12K positive COVID tests were registered with an approximately 11:1 ratio of B.1.1.7 versus B.1.617.2.
In Summary
There is still a significant level of uncertainty in the true danger of various VOCs, especially over a longer time frame. The current loose consensus seems that B.1.1.7 (Alpha) is unlikely to defeat a vaccination induced defense on its own. While B.1.351 (Beta) is significantly more dangerous than B.1.1.7 (Alpha), it still seems containable with current methods. P.1 (Gamma) is likely intermediate in threat, but more data could change this notion easily. B.1.617.2 (Delta) is clearly spreading very fast in the UK, and now also in the U.S. and other places around the world. It is suggested that the B.1.617.2 variant (Delta variant) may be the most dangerous due to seemingly much more efficient transmission than older variants like B.1.1.7 (Alpha) and the fact that unvaccinated or incompletely vaccinated persons are at significant risk from it.
References
Abu-Rabbad et al., Effectiveness of the BNT162b2 Covid-19 Vaccine against the B.1.1.7 and B.1.351 Variants. (2021) N Engl J Med., May 5;NEJMc2104974.
Allen et al., Increased household transmission of COVID-19 cases associated with SARS-CoV-2 Variant of Concern B.1.617.2: a national case- control study. (2021) Public Health England.
Bernal et al., Effectiveness of COVID-19 vaccines against the B.1.617.2 variant. (2021) medRxiv, https://doi.org/10.1101/2021.05.22.21257658.
Carvalho et al., The first 12 months of COVID-19: a timeline of immunological insights. (2021) Nat Rev Immunol, Apr;21(4):245-256.
Cele et al., Escape of SARS-CoV-2 501Y.V2 from neutralization by convalescent plasma. (2021) Nature, May;593(7857):142-146.
Dan et al., Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. (2021) Science, Feb 5;371(6529):eabf4063. doi: 10.1126/science.abf4063.
Deng et al., Transmission, infectivity, and neutralization of a spike L452R SARS-CoV-2 variant. (2021) Cell, Apr 20;S0092-8674(21)00505-5.
Faria et al., Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. (2021) Science, May 21;372(6544):815-821.
Garcia-Beltran et al., Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. (2021) Cell, Apr 29;184(9):2372-2383.
Greaney et al., Comprehensive mapping of mutations in the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human plasma antibodies. (2021) Cell Host Microbe, Mar 10;29(3):463-476.
Hoffmann et al., SARS-CoV-2 variants B.1.351 and P.1 escape from neutralizing antibodies. (2021) Cell, Apr 29;184(9):2384-2393.
Hoffmann et al., SARS-CoV-2 variant B.1.617 is resistant to Bamlanivimab and evades antibodies induced by infection and vaccination. (2021b) bioRxiv, https://doi.org/10.1101/2021.05.04.442663.
Kuchinski et al., Mutations in emerging variant of concern lineages disrupt genomic sequencing of SARS-CoV-2 clinical specimens. (2021) medRxiv, https://doi.org/10.1101/2021.06.01.21258181.
Li et al., SARS-CoV-2 501Y.V2 variants lack higher infectivity but do have immune escape. (2021) Cell, Apr 29;184(9):2362-2371.
Liu, J. et al., BNT162b2-elicited neutralization of B.1.617 and other SARS-CoV-2 variants. (2021) Nature, Jun 10. doi: 10.1038/s41586-021-03693-y.
Liu, Z. et al., Identification of SARS-CoV-2 spike mutations that attenuate monoclonal and serum antibody neutralization. (2021) Cell Host Microbe, Mar 10;29(3):477-488.
Liu, Y. et al., Neutralizing Activity of BNT162b2-Elicited Serum. (2021) N Engl J Med., Apr 15;384(15):1466-1468.
Madhi et al., Efficacy of the ChAdOx1 nCoV-19 Covid-19 Vaccine against the B.1.351 Variant. (2021) N Engl J Med., May 20;384(20):1885-1898.
McCallum et al., N-terminal domain antigenic mapping reveals a site of vulnerability for SARS-CoV-2. (2021) Cell, Apr 29;184(9):2332-2347.
McCarthy et al., Recurrent deletions in the SARS-CoV-2 spike glycoprotein drive antibody escape. (2021) Science, Mar 12;371(6534):1139-1142.
Planas et al., Sensitivity of infectious SARS-CoV-2 B.1.1.7 and B.1.351 variants to neutralizing antibodies. (2021) Nat Med., May;27(5):917-924.
Planas et al., Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. (2021b) Nature, Jul 8. doi: 10.1038/s41586-021-03777-9.
Plante et al., The variant gambit: COVID-19’s next move. (2021) Cell Host Microbe, Apr 14;29(4):508-515.
Stamatatos et al., mRNA vaccination boosts cross-variant neutralizing antibodies elicited by SARS-CoV-2 infection. (2021) Science, Mar 25;eabg9175. doi: 10.1126/science.abg9175.
Tegally et al., Detection of a SARS-CoV-2 variant of concern in South Africa. (2021) Nature, Apr;592(7854):438-443.
Thompson et al., Circulating SARS-CoV-2 spike N439K variants maintain fitness while evading antibody-mediated immunity. (2021) Cell, Mar 4;184(5):1171-1187.
Wall et al., Neutralising antibody activity against SARS-CoV-2 VOCs B.1.617.2 and B.1.351 by BNT162b2 vaccination. (2021) Lancet, Jun 3;S0140-6736(21)01290-3.
Wang et al., Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7. (2021) Nature, May;593(7857):130-135.
Wang et al., Increased resistance of SARS-CoV-2 variant P.1 to antibody neutralization. (2021b) Cell Host Microbe, May 12;29(5):747-751.
Washington et al., Emergence and rapid transmission of SARS-CoV-2 B.1.1.7 in the United States. (2021) Cell, May 13;184(10):2587-2594.
Winger and Caspari, The Spike of Concern-The Novel Variants of SARS-CoV-2. (2021) Viruses, May 27;13(6):1002.
Zhou et al., Evidence of escape of SARS-CoV-2 variant B.1.351 from natural and vaccine-induced sera. (2021) Cell, Apr 29;184(9):2348-2361.






